

 journal menu
journal menu





issue contents
September 2008 issue
Cover illustration: Crystal packing of the c6-type cytochrome OmcF from Geobacter sulfurreducens (p. 919). Packing interactions are mediated via an extended Strep-tag II and linker sequence that forms the only connection between adjacent layers of protein within the crystal (3dp5).
research papers






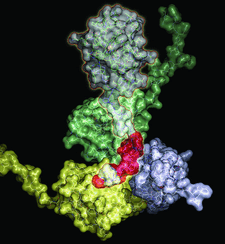
Crystal structures of ligand-free wild-type and activated zebrafish Plk1 kinase domains reveal the organization of the secondary structural elements around the active site. The cocrystal structure of wild-type Plk1 with ADP documents the hydrolysis of ATP and reveals the autophosphorylation site, while the cocrystal structure of the activated kinase domain with wortmannin, a covalent inhibitor of the PI3 kinase, shows the binding mode of the small molecule to the enzyme and could facilitate the design of more potent Plk1 inhibitors.
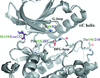
The structure of the Strep-tag II-tagged monohaem c-type cytochrome OmcF from G. sulfurreducens was determined by Fe SAD on a rotating-anode X-ray source. Packing analysis reveals that the N-terminal Strep-tag II is the exclusive mediator of crystal contacts along the c axis, providing crystal contacts to three monomers simultaneously.
PDB reference: OmcF, 3dp5, r3dp5sf
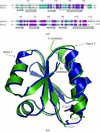
Yeast Grx3 is involved in iron-responsive transcription regulation. The single active site thiol of the thioredoxin-like domain is in a flexible surface loop as suggested by its partial disorder.
The X-ray crystal structure of a soluble Rieske ferredoxin from M. musculus was solved at 2.07 Å resolution, revealing an iron–sulfur cluster-binding domain with similar architecture to the Rieske-type domains of bacterial aromatic dioxygenases. The ferredoxin was also shown to be capable of accepting electrons from both eukaryotic and prokaryotic oxidoreductases.
PDB reference: Rieske-type ferredoxin, 3d89, r3d89sf

Crystal structures of three PCNA homologues from Sulfolobus solfataricus have been determined in different oligomeric states. The structural features in different assemblies present a molecular model for clamp-ring forming and opening.
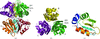
Structure of the C-terminal domain of the arginine repressor protein from Mycobacterium tuberculosis
The structure of the core domain of the arginine repressor protein from M. tuberculosis has been determined with (1.85 Å resolution) and without (2.15 Å resolution) the arginine corepressor bound. Three additional arginine molecules have been found to bind to the core domain hexamer at high (0.2 M) arginine concentration.
Open  access
access
access
High-molecular-weight poly-γ-glutamic acid-based polymers have been synthesized, tested and adopted for protein crystallization.
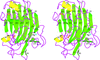
The 1.5 Å resolution crystal structure of the catalytic domain of S. sioyaensis endo-1,3-β-glucanase, a member of glycosyl hydrolase family 16, revealed an active-site cleft that was closed at one end for specific 1,3-β-glucan substrate binding.
PDB reference: endo-1,3-β-glucanase catalytic domain, 3dgt, r3dgtsf
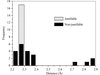
The bond distances that bind the haem group to the globin molecule in the crystallographic structures of haemoglobins deposited in the Protein Data Bank were evaluated by comparison with the same bond distance determined using EXAFS and X-ray absorption techniques.
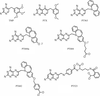
The structures of mouse DHFR holo enzyme and a ternary complex with NADPH and a potent inhibitor are described.

The new crystal structures of viscotoxins A1 and B2 have been solved by sulfur-SAD phasing and ab initio direct methods, respectively.
short communications
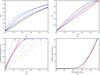
Pseudo-merohedral twinning was detected in crystals of the dihaem c-type cytochrome DHC2 from G. sulfurreducens.
letters to the editor
Free
Comment on Structural basis for cofactor-independent dioxygenation in vancomycin biosynthesis by Widboom et al. (2007), Nature (London), 447, 342–345.







