

 journal menu
journal menu





issue contents
October 2011 issue
Cover illustration: Substrate binding of a GH5 endoglucanase from the ruminal fungus Piromyces rhizinflata (Tseng et al., p. 1189).
editorial




Free 

structural communications



Metallo-β-lactamase enzymes confer resistance to a wide variety of commonly used antibiotics; this resistance poses a significant threat to human health where it arises in a pathogenic bacterium. Here, the structure of the novel metallo-β-lactamase NDM-1, which is capable of providing resistance against a broad spectrum of antibacterial agents, has been solved.
PDB reference: New Delhi metallo-β-lactamase 1, 3zr9
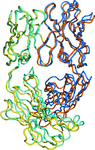
Comparison of the Fab structure in the free and antigen-bound form indicates an induced-fit mechanism of IL-13 recognition by antibody C836 through rigid-body rotation of the VL and VH domains.
PDB reference: C836 Fab, 3l7e
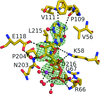
ATP-citrate lyase catalyzes the production of acetyl-CoA from citrate, CoA and ATP. Crystals were grown of the amino-terminal portion of the human enzyme in the presence of tartrate, ATP and magnesium ions. The crystal structure shows the inhibitor tartrate and the product ADP–Mg2+ bound to the protein.
PDB reference: ATP-citrate lyase, 3pff

The crystal structures of both native manganese SOD2 and iron-substituted SOD2 from S. cerevisiae were solved. The surface-potential properties showed a similar pattern to those of Cu/Zn-SODs.
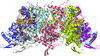
Structures of the nucleocapsid protein of human respiratory syncytial virus have been obtained from two crystal forms. The packing in one crystal form suggests that ionic interactions contribute to the stability of the native helical form of HRSVN.
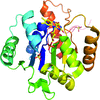
The crystal structure of recombinant UbiX has been determined to 1.5 Å resolution.
PDB reference: PA4019, 3zqu
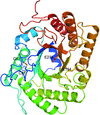
The structure of a complex of an endoglucanase with cellotriose was determined with a hexagonal unit cell and showed how the substrate interacts with the enzyme.
crystallization communications

Crystallization and preliminary X-ray diffraction studies of sortase A from Streptococcus pneumoniae
Sortase A from S. pneumoniae has been crystallized in two crystal forms: diamond-shaped (construct ΔN59SrtA) and rod-shaped (construct ΔN81SrtA). The diamond-shaped crystals diffracted poorly to 4.0 Å resolution and belonged to a tetragonal system, whereas the rod-shaped crystals diffracted to 2.91 Å resolution and belonged to space group P21.

The hexameric ND1 fragment of p97/VCP was crystallized in complex with the UBX domain of FAF1. X-ray diffraction data were collected to 3.60 Å resolution and the crystals belonged to space group I222.
Alkyl hydroperoxide reductase from Anabaena sp. PCC 7120 was expressed, purified and crystallized. Diffraction data to 2.5 Å resolution revealed that the crystals belonged to space group P212121, with unit-cell parameters a = 80, b = 102, c = 109.6 Å.

Crystals of FimP from A. oris were obtained. To facilitate selenomethionine labelling, three methionines were introduced by site-directed mutagenesis.

The complex of MoPrP(120–232) and Fab POM1 has been crystallized (space group C2, unit-cell parameters a = 83.68, b = 106.9, c = 76.25 Å, β = 95.6°). Diffraction data to 2.30 Å resolution have been collected using synchrotron radiation.

The rod domain of α-actinin-2 from E. histolytica, the aetiological agent of human amoebiasis, was crystallized and synchrotron diffraction data were collected from crystals vitrified in liquid nitrogen.

In order to confirm the effect on cocrystallization, two N-terminally truncated variants of a thermostable endoglucanase from the thermophilic bacterium Fervidobacterium nodosum Rt17-B1 were constructed, purified and cocrystallized at 291 K.

Crystals of betaine aldehyde dehydrogenase 2 from rice (O. sativa L.) belonged to a C-centred orthorhombic space group and diffraceted X-rays to 2.6 Å resolution.

The 2AB protein derived from the nonstructural P2 region of hepatitis A virus has been cloned, purified and crystallized. The preliminary characterization of native and selenomethionine-derivative crystals is reported.

ChuZ–haemin complex crystals belonged to space group C2221, with unit-cell parameters a = 106.474, b = 106.698, c = 52.464 Å, α = β = γ = 90°. Diffraction data have been collected to 2.4 Å resolution.

A truncated heterodimer of human PSPC1–NONO protein, a paraspeckle-specific complex, has been crystallized and the diffraction data collected to a resolution of 1.9 Å.

The DHH-family phosphoesterase NrnA is involved in the degradation of small RNA oligomers. B. subtilis NrnA crystallized in space group P21 and diffracted X-rays to a resolution of 2.0 Å.
Yeast Sfh3, a Sec14-like phosphatidylinositol-transfer protein with unique functional properties, has been crystallized. X-ray data sets were collected from native and selenomethionine-substituted crystals to 2.2 and 1.93 Å resolution, respectively.

The haemagglutinin subcomponent HA3 of the type B botulinum neurotoxin complex, which is important in toxin absorption from the gastrointestinal tract, has been expressed, purified and subsequently crystallized in two crystal forms at different pH values.

The crystallization of PBP4 from L. monocytogenes is reported.
Open access
access
The C-terminal cytosolic domain of mouse anoctamin 1 (mANO1, also known as TMEM16A) was cloned, overexpressed, purified and crystallized. The crystals belonged to space group P212121 and diffracted to 2.3 Å resolution.

The crystallization of the TLDc domain of oxidation resistance protein 2 from zebrafish is reported.

The successful preparation of a mutant KH domain representing the first KH domain of PCBP1 and its crystallization in complex with a C-rich DNA are reported. This structure is anticipated to provide high-resolution information that will allow better understanding of the basis of cytosine specificity by PCBPs.

To further understand the structural basis of MmeI (a type IIL restriction enzyme) DNA-binding specificity, the enzyme has been crystallized in complex with its DNA substrate.

eCGP123, an extremely stable GFP with photoswitching properties, has been expressed, purified and crystallized. A diffraction data set has been collected at 2.10 Å resolution.

The gene for 3-ketosteroid Δ4-(5α)-dehydrogenase from R. jostii RHA1 was cloned and overexpressed in E. coli and the protein product was purified and crystallized using the hanging-drop vapour-diffusion method. The crystals belonged to space group C2221 and diffraction data were collected to a resolution of 1.6 Å.

The C-terminal domain of FbiB, a bifunctional protein that is essential for the biosynthesis of cofactor F420 in M. tuberculosis, has been expressed, purified and crystallized. The crystals diffracted to 2.0 Å resolution and were suitable for structure determination.

PaaX, the main regulator of the phenylacetic acid degradation pathway in bacteria, has been crystallized using the sitting-drop vapour-diffusion method. Preliminary X-ray data to 2.3 Å resolution allowed determination of its unit-cell parameters and space group and initiation of its structure determination.
Recombinant L-ribose isomerase from Acinetobacter sp. was crystallized. Diffraction data were collected to 2.2 Å resolution.

Phosphoglycerate kinase, a two-domain enzyme involved in the glycolytic pathway, has been crystallized. Both the N-terminal domain and a ternary complex of the full-length enzyme with substrates were crystallized by the sitting-drop vapour-diffusion method. Inclusion of the substrates was critical in order to obtain crystals of the full-length protein.

Methods that led to the successful crystallization of TLR3 ECD in complex with Fab fragments from three noncompeting, neutralizing anti-TLR3 antibodies are presented. The crystallization of this 238 kDa complex was achieved through fine purification of the quaternary complex of TLR3 ECD with three Fab fragments combined with microseed matrix screening and additive screening.

Methods for the expression, purification and crystallization of the class C acid phosphatase from M. bovis are reported. This enzyme is atypical in that it is nearly 20 kDa larger than other known class C acid phosphatases.
laboratory communications
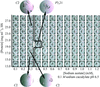
Protein crystals typically deteriorate upon soaking in solutions very different from those used to grow them (the `mother liquor'). This is particularly true of solutions containing lower salt concentrations and organic solvents (i.e. DMSO, ethanol). Optimization of the relative humidity environment of the crystals can compensate for these effects and allow recovery of diffraction quality.
![[publBio]](/logos/publbio.gif)




